臨床生物化學/載脂蛋白的基因結構及表型
| 醫(yī)學電子書 >> 《臨床生物化學》 >> 血漿蛋白及其代謝紊亂 >> 載脂蛋白 >> 載脂蛋白的基因結構及表型 |
| 臨床生物化學 |
|
|
|
各種生物都能通過生殖產(chǎn)生子代,子代和親代之間,不論在形態(tài)構造或生理功能的特點上都很相似,這種現(xiàn)象稱為遺傳(heredity)。但是,親代和子代之間,子代的各個體之間不會完全相同,總會有所差異,這種現(xiàn)象叫變異(variation)。遺傳和變異是生命的特征。遺傳和變異的現(xiàn)象是多樣而復雜的,正因為如此,才導致生物界的多種多樣性,生物體所具有的遺傳性狀稱為表型或表現(xiàn)型(phernotype)。生物體所具有的特異基因成分稱為基因型(genotype)。表型是基因型與環(huán)境因素相互作用的結果。遺傳物質是相對穩(wěn)定的,但是又是可變的,遺傳物質的變化以及由其所引起表型的改變,稱為突變(mutation)。遺傳物質突變包括染色體畸變和基因突變。基因突變是染色體中某一點上發(fā)生化學改變,所以又稱為點突變(pointmutation)。基因結構和遺傳表型的研究是深入了解脂蛋白代謝缺陷癥的分子生物學基礎,逆向遺傳學方法(reversegeneticapproach)則使其有可能在蛋白質水平系統(tǒng)地分析結構和功能的關系。現(xiàn)已采用一個特定的cDNA探針從基因文庫中篩選所需要的基因進行cDNA克隆,測定其核苷酸序列,然后從核苷酸序列推斷蛋白質氨基酸序列。目前,已分離出許多與動脈粥樣硬化有關的脂蛋白的cDNA克隆,并將其蛋白質一級結構的氨基酸排列順序和基因的核苷酸順序測出。現(xiàn)已查明,ApoAⅠ、AⅣ、E、B、CⅡ和(a)都存在著異構體,也就是說存在著各種不同的表型或基因型,并可分別從蛋白質水平和核酸水平進行分型。現(xiàn)分別介紹幾種主要載脂蛋白的基因結構。
(二)載脂蛋白基因結構特點
人血漿中載脂蛋白的結構及功能,經(jīng)過近十年的深入研究,已了解得較為清楚。大部分載脂蛋白的基因和cDNA都已得到分離和確定,其核苷酸順序也進行了測定。除ApoAⅣ,B、(a)外,它們的共同特點是含有三個內含子(intron)和四個外顯子(exon),其內含子插入外顯子的位置大致相同,基本上按照生理功能的不同,將其加以分隔。第一個內含子把5′-末端的非翻譯區(qū)和翻譯區(qū)分開;第二個內含子把信號肽編碼(singnalpeptide)和功能蛋白編碼區(qū)分開;第三個內含子則把原肽編碼區(qū)和成熟肽編碼區(qū)分開。這些基因的第一、二、三外顯子的核苷酸數(shù)量也相差無幾,第四個外顯子核苷酸數(shù)量不同而導致各種載脂蛋白基因長度不同。從生物進化角度考慮,上述載脂蛋白基因結構相似性,提示可能來源于一個共同的祖先,即ApoCⅠ基因。ApoAⅣ與其他載脂蛋白基因結構不同,它只含有三個外顯子。載脂蛋白基因結構的另一特點是幾個基因相接很近,定位于同一染色體的一個位點上或附近,呈緊密連鎖狀態(tài)。如ApoAⅠ、CⅢ和AⅣ基因位于第11號染色體長臂2區(qū),形成一個約15kb的基因簇。還有一個緊密連鎖的基因簇是ApoE、CⅠ和CⅡ基因,同位于第19號染色體長臂3區(qū),見圖4-4。
ApoA-Ⅱ基因定位于第1號染色體長臂2區(qū),ApoB基因定位于第2號染色體短臂2區(qū),Apo(a)基因定位于第6號染色體長臂2區(qū)。
(三)載脂蛋白基因結構
1.ApoAⅠApoAⅠ基因長1863bp,含有三個內含子,第一個內含子位于5′端非翻譯區(qū);第二個內含子位于翻譯區(qū)的AⅠ前肽區(qū)內;第三個內含子插入翻譯成熟AⅠ第43氨基酸殘基處。ApoAⅠ基因含有四個外顯子,分布于ApoAⅠ基因的不同區(qū)域,ApoAⅠ基因與ApoCⅢ、AⅣ基因相連成簇,CⅢ基因居中,轉錄方向與AⅠ和AⅣ基因相反。位于AⅠ和CⅢ基因共同3′區(qū)的DNA序列,可能參與對AⅠ基因的轉錄調控。
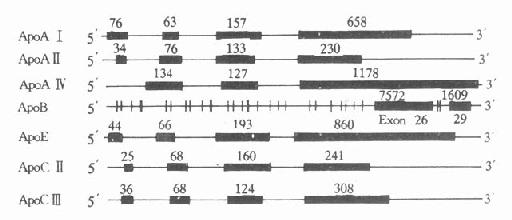
圖4-4 人載脂蛋白AⅠ、AⅡ、AⅣ、B100、
CⅡ、CⅢ和E基因結構示意圖
粗線代表外顯子,粗線之間的細線代表內含子,粗線上緣數(shù)字
代表該段核苷酸數(shù)目
2.ApoBApoB族位于2號染色體P23→Pter區(qū),是由非翻譯區(qū)、編碼區(qū)、TAA終止密碼子和一個3′端的非翻譯區(qū)組成。ApoB100基因全長43kb,含29個外顯子和28個內含子見圖4-4,其中第26和第29兩個外顯子特別長,分別含有7552和1905bp,外顯子2最短,僅39bp(從211-249)。內含子則以第27個為最短(107bp)。人群中至少有14種不同的3′端高變異等位基因區(qū),75%的人群在此區(qū)是不均一的。
ApoB48和ApoB100除了在結構上有關外,ApoB48的形成機制目前尚無完全一致的看法,主要認為有合成ApoB48的基因存在。1987年被發(fā)現(xiàn)ApoB48是由ApoB100通過一種新的機制涉及到mRNA的編輯而產(chǎn)生的。在測定從人小腸基因庫分離的ApoBcDNA的序列時發(fā)現(xiàn),小腸ApoBcDNA的第6666個核苷酸為T,而從肝分離的ApoBcDNA克隆在此位置為C。將T替換C則6666處產(chǎn)生一終止編碼(TAA),TAA替換CAA編碼使ApoB100的2153位氨基酸應為Gln,預示血漿中存在的ApoB48應是相當于ApoB100的2153氨基末端為Gln。這一預測后來得到實驗證實,并發(fā)現(xiàn)核苷酸上6666的替換C→T只發(fā)生在小腸的mRNA上,而不發(fā)生在小腸基因組(genomic)DNA上,因此這是轉錄以后的一種特殊形成的編輯小腸mRNA的結果.
3.ApoE人ApoE基因位于19號染色體長臂3區(qū),含有四個外顯子和三個內含子。
1975年首先觀察到ApoE的多態(tài)性,利用等電聚焦電泳和SDS-PAGE可以確認ApoE的多態(tài)性。實驗表明,ApoE有三種異構體(isoform)即E2、E3和E4。有的人只含有一種主要異構體即純合子,有的人可含二種主要異構體為雜合子。由此可見,人群中可有六種不同的表現(xiàn)。根據(jù)ApoE表型提出ApoE基因模型認為,ApoE的合成是由位于一個基因位點上的三個等位基因所控制,即E2、E3和E4,每一個等位基因對應于一個主要異構體,產(chǎn)生三種純合子(E2/2,E3/3,E4/4)和三種雜合子(E2/3,E2/4,E3/4)共六種常見表型,另外,還有極少見的異構體。一般認為,次要異構體是由主要異構體翻譯后,經(jīng)唾液酸糖化修飾后轉變而來。ApoE3/3型又稱野生型。ApoE的基因序列的112位和158位兩種氨基酸殘基即精氨酸(Arg)和半胱氨酸(Cys)的交換決定了異構體的種類。ApoE4在這兩個位置上都是Arg;E2都是Cys;112和158位是Arg者為ApoE3異構體。自然人群中,基因頻率(3)分布最高,ApoE3/3表型分布約70%,見圖4-5。
4.ApoC族ApoCⅡ基因有3347bp,含有4個外顯子和3個內含子。ApoCⅡ的羧基末端氨基酸序列是激活脂蛋白脂肪酶的活性功能區(qū)域。ApoCⅢ基因含有3133bp,有4個外顯子和3個內含子。

圖4-5 人ApoE三種主要異構體的氨基酸殘基及基因密碼的改變位置
5.Apo(a)運用cDNA探針進行染色體定位研究時發(fā)現(xiàn),Apo(a)的基因位點在人第6號染色體長臂2區(qū)6-7帶間,與血纖溶酶原(PLG)的基因位點有部分重疊。測定PLG基因跨距為525kb,由18個內含子與19個外顯子組成,5個Kringle結構由各自兩個外顯子編碼。Apo(a)cDNA分析表明,Apo(a)與PLG的基因有很多相似之處。
通過家系研究,目前已發(fā)現(xiàn)Apo(a)基因位點中至少有26個等位基因與多態(tài)性有關。這些等位基因至少表達有34種Apo(a)異構體。
| 關于“臨床生物化學/載脂蛋白的基因結構及表型”的留言: | |
|
目前暫無留言 | |
| 添加留言 | |